Introduction
The NEURO-M Predictive Temperament Assessment has proven to be highly reliable in workplace applications, most often as an aid for recruitment, career planning and personal development. In particular, it has been especially useful in predicting behavioural response patterns to stress that seem “out-of-character”, and often sit outside of an individual’s preferred view of themselves.
This paper describes the ancient origins and neurobiological underpinnings of temperament and temperament change, linking earlier ideas based on observations and flawed sense-making to modern scientific research on brain design and function. This has led to the development of a better-informed and more reliable and useful understanding and interpretation of temperament assessment results.
An overview to the concept of temperament typologies
The four basic temperament types described in various temperament-typing mechanisms are a modern-commercial development of the ancient concept of “humorism”, where chemical compounds within the blood were believed to be at the heart of human behaviours and behavioural patterns.
Hippocrates (c460 – c370 BC) observed that the blood of unhealthy people, rather than simply remaining a consistent red, quickly formed layers, a process today explained by erythrocyte sedimentation (1) where in a vial of blood a yellow layer of serum tops a grey clot, with the red blood cells below that and the blackish de-oxygenated hemoglobin at the base. Hippocrates described these four “humors” (humor is a translation of the Greek word Chymos, meaning juice or sap), seeing the yellow layer as “bile” (chole), the grey resembling mucus (phlegm), the red blood (sanguis, from the Latin), and the almost-black bottom layer as black bile (melancholia)(2). The concept of four humors also fitted the prevailing concepts of “fours” as a naturally occurring phenomenon in elements, winds, seasons and qualities.
Hippocrates speculated that imbalances between humors could be seen through imbalances in those humors, and/or a failure for them to blend, and this became a significant, albeit incorrect, influence on medicine for the next two millennia (linked to blood-letting and other “cures”)(3).
Galen (4), Avicenna, Culpepper and others including Kant, Steiner, Adler and Fromm drew on and interpreted these ideas over time in an attempt to understand and explain behavioural traits, the commonalities between them and an origin for them. Certainly, the behavioural patterns observed throughout centuries appeared consistent, and so it seems likely that there was a rational explanation for them. Specifically, Galan ascribed moods, emotions and behaviours to the proportions the humors were mixed, and named the four temperament (from the Latin “tempere”, meaning “to mix”) types according to the dominant humor associated with them. He associated a phlegmatic temperament with disinterested, dullness and low energy, a sanguine temperament with passion, laughter and fun, a melancholy temperament with being gloomy and depressive, and a choleric temperament with being short-tempered and fiery (3, 5).
Example attributes of those four temperament types, or trait-groupings, as broadly interpreted now, include:
- Sanguine: Naturally outgoing, full of ideas, highly social and socially confident, popular, light-hearted, story and joke-teller, animated, fun-loving, impulsive, not good with money or great with details, persuasive. Birthdays are another reason to get together and have fun! Unlikely to procrastinate socially but can often fail to see tasks or ideas through.
- Choleric: Naturally strong-willed, socially dominant, competitive, outcome and big-picture focused, stubborn, belligerent, efficacious, leader, determined, capable of making hard choices, driven, seeks to control or draw on others to succeed. Unlikely to procrastinate. Own birthday is important, recognises that other birthdays matters to those people.
- Phlegmatic: Naturally relationship-focused, peacemaker, supporter, servant, compliant, non-assertive, can lack focus and drive, will strive to not let others down, more likely to withdraw from conflict or upset. Can procrastinate through worry of impacts on others, or lack of confidence. Remembers everyone else’s birthdays – and organises the cake.
- Melancholy: Naturally task and details-focused. Likely not great with emotional awareness, comfort expression and interpersonal skills. May spend too long overdoing tasks. Conscious thoughts are rational, logical, researched, reviewed, contemplated, checked and rechecked. Technically artistic. Sees complexity and tries to account for it, can lead to procrastination. Unlikely to remember birthdays – but may create a system to remember them.
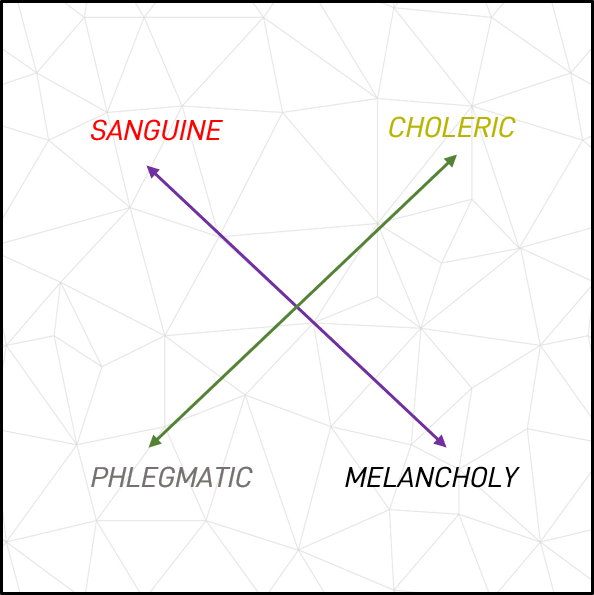 Figure 1. Galen’s Humors demonstrating opposites
Figure 1. Galen’s Humors demonstrating opposites
Today’s popular DiSC® personality types and profiling business model is based on W. M. Marston’s modern western version of the ideas of Galan and Hippocrates, albeit with very little recognition of its origins, with validity referenced as longitudinal stability and various other measures. Longitudinal stability is key to useful and reliable outcomes for assessing personality and temperament (rather than mood or positivity), and it is a strength for DiSC, the Five Factor Model of Personality and some others, but certainly not all (notably where initial response simplification is followed by specific interpretation). Unfortunately, the lack of neurobiological grounding inhibits the potential accuracy and usefulness of most of these types of assessments.
It is one thing to notice patterns and then to estimate future accuracy through past correlation. It is another to start with the cause before arriving at an effect, especially as simplification and categorisation can have significant consequences for recruitment, career planning, self-development, personal identity and, ultimately, well-being.
The neurobiology of temperament types
The concept of trait commonality and predictability associated with the general concept of temperament typologies has proven popular and somewhat reliable, despite deficits in why, or objective accuracy. To improve our basic understanding of what a temperament is, how it forms and why it is relatively stable for adults. The brain, as with any other body part, first grows according to instructions coded in DNA, and subject to nutrition, chemical influences, health of the mother and other environmental factors (6-8).
At birth the parts of the brain essential for life are fully formed so that basic functions (breathing, heartbeat, digestion etc) can occur and the baby can survive. The powerful emotional centre (the so-called limbic region), is only partially formed, and the region involved with intelligence, emotional control and rational learning (the neocortex) is quite immature. By about the age of three, the limbic region is functional, but it takes a further 22 or so years for the neocortex to fully mature, albeit not lineally.
Like muscle groups and bones, the brain’s development is influenced by experiences in both the short and long terms. And like muscles in particular, it responds to the demands it perceives are placed upon it to best help its host cope, survive and reproduce. The first three years of life in particular, where experiences shape the development of the powerful limbic region, affect how genes are expressed and synaptic pathways are connected, involving basic brain processes of growth, development and neuroplasticity, being differentiation (the conversion of stem cells into specialised neurons located within different brain regions and with differing capabilities and functions), epigenetics (the expression of genes according to experiences, environments and nutrition), synaptogenesis (the creation of connections between neurons) and myelinogenesis (the formation of a protective and strengthening sheath over synaptic connections) (9). These processes dictate that the brain will evolve as a result of both DNA and experiences, and responds (and continues to change) according to external and internal stimuli and nutrition.
For this reason, it is apparent that the foundation for temperament is heavily influenced in these first years of brain growth and development, most heavily influenced by the DNA and the environment provided by parents (10). As children go to school, their brains become influenced by new social and hierarchical environments, most often quickly learning (creating new synaptic connections) and adapting behaviours (creating and following new or modified synaptic pathways), influencing and shaping personality along the way as the child learns how to survive unfamiliar, complex and at times unsafe social situations.
The transition to adulthood, triggered by large hormone releases and continually influenced by social environments is another time of accelerated brain change, seeing, at first, changes in traits, and then gradual stabilisation (11) with increases in conscientiousness, agreeableness and emotional stability (12). Notably, similar changes are likely to be occurring in the brains of the social group (typically other schoolchildren), seeing this as much a social evolution as a personal one and often including the beginning of separation from parents in the search for independence and identity (13), a potentially traumatic and confusing time for families.
As the neocortex continues to full maturity at around 25 years of age, stabilisation continues and change continues to slow, and at that stage it might be said that one’s temperament is fully formed. But… it isn’t. Through the four basic processes described earlier, along with injury, disease, atrophy and changes in hormones and nutrition, the brain continues to change in response to its experiences, albeit at a relatively slow rate. However, the impacts of severe emotional influences, such as trauma (e.g. PTSD), along with other continual adaptations to specific people, events and environments (potentially including therapy and mental self-care), sees the brain changing more rapidly in stress-related (limbic-based) responses. (Changes in the neocortex also occur throughout life and can occur quickly as rational learning and skills development but are usually less influential in trait formation unless an individual is deeply affected by them, and/or consciously and diligently chooses to exercise them – meditation is an example of this.) However, the extra effort required for thinking and behaving “contra-trait” (based mostly on synaptogenesis and myelinogenesis) generally sees stable, habitual “trait-typical” actions dominate behaviours (14).
From this we can see that a temperament, being a blend of behavioural traits arising from the creation and strengthening of synaptic pathways (15) as habitual responses, is highly influenced by DNA and early life, is heavily influenced by social change through childhood, and then is again heavily influenced by the hormonal cascade of puberty before slowing and stabilising into the early twenties, thereafter changing (most likely consolidation) through experiences at a much slower rate (16). At this stage, an individual will have developed a set of behavioural habits (the strongest and fastest default synaptic pathways) that allow the person to feel relatively safe, with some stability and security in their relationships and personal identity, and how they interact and cope socially in feeling a sense of control in their life (i.e., satisfying basic needs). It is now likely that the behavioural habits (or, collectively, traits) are reliably measurable, and consistencies in and between behavioural patterns can be perceived as personality (a complex construct) or classified as temperaments (a simplifying and generalising construct).
The Origins of the NEURO-M Predictive Temperament Assessment
Self-assessments, including temperament assessments, have inherent limitations in design and interpretation. Hippocrates typology, DiSC®, Myers-Briggs, the Five Factor Model of Personality, The Herrmann Brain Dominance Instrument and others present common behaviours in ways that allowed people to feel they could “understand” themselves and/or categorise others (simplification, stereotyping and generalisations appeal to the basic need for orientation (17)). Personal assessments and/or psychological diagnosis can lead to attaching one’s own behaviours to identity (a powerful basic need linked to self-esteem), which then sees justification and/or repetition of behaviours as reinforcement or enhancement of that basic need. This supports a positive-feedback loop that can see a narrowing and strengthening of behaviours consistent with identity, whether or not they are helpful for the individual or others. Where an individual chooses not to “be that person” in relation to some behaviours, or “be that person” in regard to others, quite helpful behavioural change can occur with persistence and positive reinforcement – surely this is the ultimate benefit of this type of information.
The credibility for temperament assessments lies in their ability to, having information on some common behaviours, predict other complementary behaviours. For example, an “introvert” is less likely to be as decisive or controlling as an “extrovert”; someone who is very focused and driven is also more likely to be arrogant or uncaring about the feelings of others; and perhaps someone who is big on minor details is also not likely to be the most popular or talented conversationalist. The causes of these commonalities and the predictability of temperaments (trait stability) are rarely questioned.
There are some problems with simplifying temperaments or personalities, or of the foundations for them. In particular, the idea that any person could be defined as a single point on a chart, or a set of letters, seemed overly and likely unhelpfully simplistic, and although such singular trait-oriented ideas can fit with the basics of genetics and brain development or theories of constructivism (being modernist and post-modernist views of essentially the same thing), it doesn’t fit with the strongest and fastest neurobiological response – the fight/flight/freeze (FFF) response system (17).
The FFF response mechanism is part of the sympathetic nervous system, which in turn is part of the autonomic nervous system, which regulates many of the body’s actions unconsciously, operating independently of conscious thought or intent. The sympathetic nervous system is always active due to its role in homeostasis, as is the parasympathetic nervous system. Each acts as a counterbalance, and at times, antidote to the other, with the sympathetic nervous system being response and action-oriented, increasing impulsivity, reactivity and simple/fast-decision-making; and the parasympathetic nervous being the opposite, returning energy to digestion, immune system, general health and slower, fuller, more complex thinking. (Reinforcement Sensitivity Theory (18) proposes a separate moderating process its authors identify as the behavioural inhibition system (BIS), but there is no evidence that such a distinct and separate system exists.)
The FFFS is described in some detail in other articles here and elsewhere, fundamentally involving a strong activation of the sympathetic nervous system, with a substantial limiting of “normal” (higher order, neocortical) brain functions (including rational thinking and emotional control) and circuitry to focus energy and attention on surviving an immediate life-threatening (as primally or reflexively perceived) crisis. For temperament typologies, this, theoretically, raises questions around whether an individual who was neatly classified as a dot on a graph acting in accordance with a given typology blend when in one state of mind (notably the state of mind when completing the questionnaire, including self-assessing past behavioural habits), might act according to a different temperament mix when there is a change, especially under significant pressure. This change contradicts the assumption of assessments that associate individuals with a single behavioural pattern (albeit across more than a single domain of temperament classification) with commensurately predictable upsides and downsides.
In addition to the neurobiology of stress responses, there were three other influences behind reviewing the concept of fixed and stable temperaments. One is continual brain change, as discussed earlier, that allows for changes in temperament, especially with trauma or focused effort. The second is the influence of environment and context – behaviour is a response. The third was an acknowledgement of the post-modernist (19) philosophical rejection of traits, where rather than a single truth or view, multiple truths exist between and within individuals, and so a trait or behavioural response in the eyes of one person might not be a trait or the same behavioural response in the eyes of another, and is in any case not attached to an individual – it may simply be a momentarily or selectively adopted or perceived response pattern. This less-judgemental type of thinking is consistent with attribution theory (20, 21), recognising that behaviours are commonly attributed by their performer as a response to an external stimulus, whereas others view them as intrinsic to the individual.
Despite interpretations and questions around causality and significance of individual behaviours, the evidence behind trait consistency and personality, both as observed behaviour and as a neurobiological inevitability is strong(14). However, the other views are a good reminder of what we don’t, and may never know, inviting the avoidance of hubris and over-simplification in what might be “known”, interpreted or assumed from an assessment, enhancing tolerance and understanding for behavioural patterns that don’t fit a reviewer’s personal sense-making, beliefs and experiences. This humble, partially-informed, curious position informs how any single assessment taken at a single point in time might be most realistically and helpfully interpreted.
Principles of The NEURO-M Predictive Temperament Assessment
Extroversion and Introversion – The NEURO-M Predictive Temperament Assessment explores opposing dimensions of two fundamental behavioural characteristics which can be associated with influential neural mechanisms. One of these is introversion versus extroversion, where the definitions of both relate to motivation schemas of approach and avoidance (17, 22) in response to the reward/loss sum (23-25) attached to interpersonal activities. Simply, those who perceive taking social risks as more often associated with gain are more likely to be confident and proactive in how they interact in socially insecure situations (e.g. with strangers, in groups) – they perceive it as an opportunity for gain (as estimated by the mesolimbic dopamine pathway), and so adopt an “approach” (gain-oriented, likely to be linked with parasympathetic nervous system up-regulation) schema. Conversely, those who more commonly associate interacting with strangers or taking an active social role with risk (mostly to self-esteem/identity), commonly adopt an avoidance schema (loss-averse, likely to be linked to sympathetic nervous system activation), seeing them typically minimise interaction and potential for social competition or conflict be avoiding the interaction, behaviours linked to introversion.
Most people fluctuate between behaviours identified as one or the other (omniversion) simply to function socially, according to how their subconscious motivation valuation network judges anticipated reward, loss, risk and effort for each situation. In most cases seeing approach (extroversion-orientation) or avoidance (introversion-orientation) dominate as a “default” habit which may remain largely similar or change in response to stressed and non-stressed situations. (The single categorisation of individuals as one or the other, even based on a 51% to 49% score, is a major flaw in, and limitation for, the MBTI, as is its philosophical but quite impractical re-interpretation of each as energy sources, rather than behavioural patterns.)
Under stress, as part of a fight or flight response at a minor or major level, the reward/risk sum attached to interacting, either through social (especially conflict) withdrawal (flight response) or increased assertiveness (fight response), may change in accordance to the perceived value of a loss – for example a normally quiet parent is not likely to simply allow a child to be harmed – the risk of temporary social pain is valued lower than the risk of permanent and extreme attachment violation. Myelinogenesis dictates that this behaviour is likely to follow a familiar response pattern in itself, but it may be quite different to avoidance responses for other, less crucial potential losses, irrespective of dominant typology.
Socio-emotional and Task Focus – The other dimension is that of task versus relationship (or socio-emotional) orientation. The neural underpinning for this is also a diametric one based on the dominance in individuals of the task positive network (TPN) or the default mode network (DMN), as differentiated through comparative deactivation (broadly consistent with alternate definitions of activation and deactivation) (26).
The DMN has two primary circuits – one is involved with thinking about the mental states of self and others (but not broader “social cognition”), with the other circuit being associated with autobiographical memory, cognitive representations of emotion, value/reward estimation, emotional self-regulation and autonomic processing. This sees the DMN involved in perceptual information, especially including ideation, emotional awareness and self-regulation. Dominant DMN activation sees individuals highly influenced by socio-emotional influences and thinking, while perhaps not being quite so good at focusing on details, purely objective rationality and task completion.
The TPN is specifically activated by non-social tasks (which simultaneously deactivate the DMN), acting to focus attention, facilitate working memory (involved in concentration), logical consideration, mathematical and scientific deductions and causal and/or mechanical reasoning. However, the TPN also reduces the influence of ethics and morality (as such things are not “rational”), and while it supports rational problem-solving, it also limits inspirational innovations which can come from unfocused, broader and more contemplative (i.e., DMN-dominant) thinking. TPN domination sees individuals concentrating (perhaps experiencing feelings of “flow” (27)), drawing on dry cognitive rationality to solve problems and achieve outcomes, while often devaluing human and social elements, including the emotions of others of the values and ethics of individuals or the social group.
This “opposing domains hypothesis” of the DMN and TPN, and its implications have been explored using fMRI to record brain activation patterns between mechanical/analytical and social tasks (28), finding that regions associated with mechanical reasoning (e.g. superior frontal sulcus, lateral PFC, intraparietal sulcus) were turned off during social tasks, and that they were active while other regions which were active for social tasks (e.g. medial parietal/posterior cingulate, medial PFC and right tempo-parietal junction) were switched off during non-social tasks. They concluded that this anti-correlation “reflects a powerful human tendency to differentiate between conscious persons and inanimate objects in both our attitudes and modes of interaction” (27 – p.396).
It was also shown (29) that objectifying and dehumanising was linked to TPN activation, indicating that people make cognitive choices when thinking about others. This can be seen in everyday work situations, where employees can be seen, or feel to be perceived, as simply a job description or a payroll number. The DMN and TPN can work together, at least partially, but generally ineffectively, in an attempt to blend, say, empathy and logical reasoning (30), being seen as insincere and manipulative. Clearly, it seems more effective and more authentic to switch between networks than to attempt to straddle both. Interestingly, there is also some evidence that the TPN is more closely linked to the sympathetic nervous system, while the DMN is linked more with the parasympathetic nervous system , suggesting that the TPN is more likely to activate defensive (avoidance) responses (31). From there it might be hypothesized that people who spend more time using their TPN are more likely to experience anxiety and reductions in cortical capacity – things that act to both increase emotional activation and work against their ability to function most effectively in that mode.
In a practical sense, both networks are vital. Just as DMN thinking can have little focus or outcome, TPN thinking can also limit problem-solving by getting stuck in repeated, narrow neural loops – for example, “sleeping on it” or intentionally not thinking about an answer, which reduces activity in a specific dead-ended synaptic pathway or loop, can allow individuals to suddenly perceive the answer they were looking for, illustrating the benefit of shifting between both networks.
People continually switch between these two modes (32), often shifting in and out of task-oriented concentration and general emotional or intuitive awareness several times a minute. As with introversion and extroversion, the neuroplasticity (including creating and strengthening specific synaptic pathways) that create habitual ways of thinking, feeling and responding see some people favouring the DMN more often at work, enabling them to connect emotionally but perhaps not get as much “work” done, while others more commonly use the TPN, focusing on tasks but perhaps failing to connect with others to gain their support (rational and emotional). For ease of use, the NEURO-M Predictive Temperament Assessment describes the influence of these two networks as task-focused (TPN-bias) and socio-emotional (DMN-bias).
Other neurobiological concepts not explored here, such as left vs right processing dominance, often presented as specific vs general, or rational vs emotional bias; and neocortical versus limbic/reptilian brain regionality (a slightly outdated concept), linked with emotional/intuitive influence versus cognitive/rational influence (a highly relevant and influential concept that directly relates to processing speed and bias), are consistent with this type of socio-emotional versus task-focused distinction. Due to the brain’s continual search for consistency (17) (compatibility in neural processing) which sees these processes continually driven to reconciliation (with varying degrees of success), DMN vs TPN activation and deactivation can be more cleanly identified.
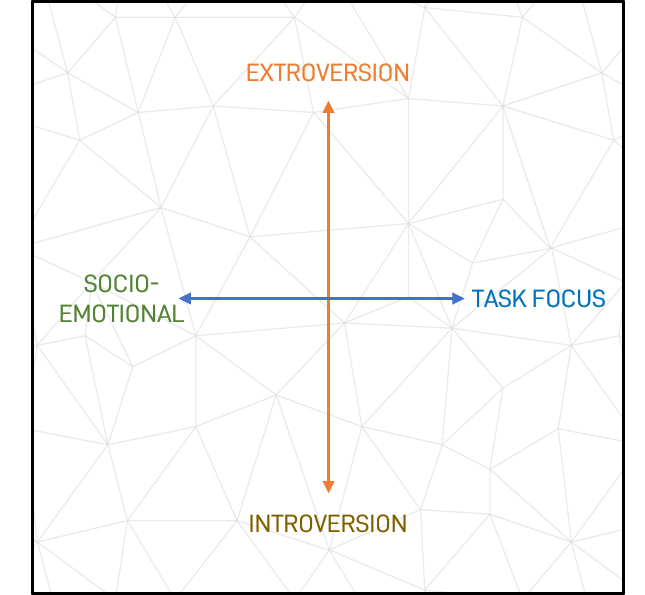 Figure 2: Opposing Basic Behavioural Properties
Figure 2: Opposing Basic Behavioural Properties
Stability –The stability of behavioural patterns is a consequence of synaptogenesis, being the creation of connections between brain cells, and myelinogenesis, the process of strengthening, protecting and increasing the speed (by up to 3000x) of commonly used connections. As a brain function, following well-myelinated pathways is also known, and therefore safer, than alternatives, irrespective of objective merit. The known, safer path comes with a little dopamine hit that, all things being equal, is not present with lesser-known “unsafe” options. In this way, experiences and responses form, reinforce and strengthen each other through life.
A Causal Explanation for Temperament Composition – Returning to the origins of temperament classifications, these opposing, and neurobiologically-founded behavioural pattern dimensions of social risk vs social reward and task-focus vs socio-emotional orientation correlate with the gaps between dimensions proposed by Galan. For example:
- Sanguine traits combine extroversion over introversion, and socio-emotional over task-focus
- Choleric traits combine extroversion over introversion, and task-focus over socio-emotional influence
- Phlegmatic traits combine introversion over extroversion, and socio-emotional over task-focus
- Melancholy traits combine introversion over extroversion, and task focus over socio-emotional influence.
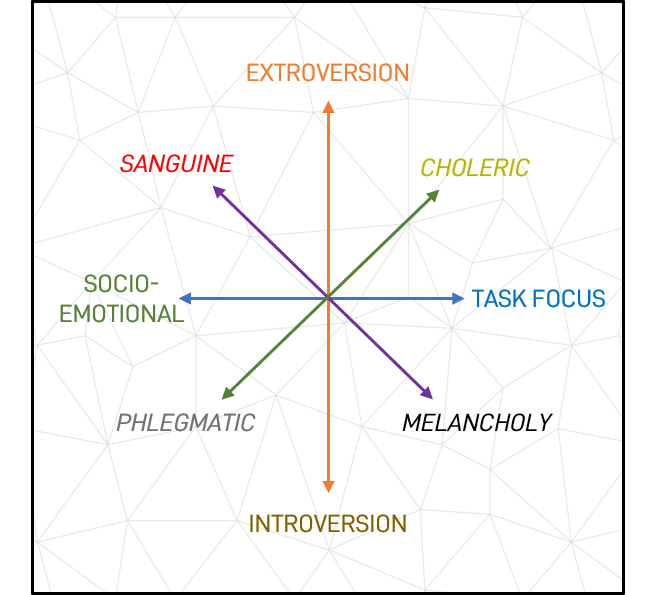 Figure 3: The complementary relationship between Galen’s Humors and Basic Behavioural Properties
Figure 3: The complementary relationship between Galen’s Humors and Basic Behavioural Properties
In this way, the NEURO-M process can both identify generally accepted trait patterns of this type of approach while also attaching them to a neurobiological base.
Research
Based on the neurobiology of the stress response, it was hypothesised that significant change in behaviours, and therefore temperament, would occur under stress in tandem with sympathetic nervous system activation, especially where FFF responses were triggered by severe fear or distress.
Data from the NEURO-M Predictive Temperament Assessment consists of 40 questions relating to common feelings and choices. 20 questions have only “positive” answer options representing “positive” behaviours reflecting a likely unstressed, parasympathetic nervous system-dominant neurobiological state. The other 20 questions offer only “negative” answer options representing “negative” behaviours likely derived from heavy sympathetic nervous system dominance, especially FFF system activation. In this way, questions specifically explored temperament with a “normal” system balance (likely high in serotonin and dopamine / dopamine anticipation and hippocampus-PFC activity) and gain-oriented (approach) motivation in comparison to a “fear” system (likely lower in serotonin and dopamine, and higher in cortisol, accompanied by higher levels of action-oriented epinephrine and norepinephrine in the body, with reduced hippocampus/PFC (rational) capacity) due to the dominance of loss-prevention (avoidance) motivation.
By only allowing participants to choose between 4 positive, or 4 negative, responses, they could not bias responses toward presenting or preserving an overly favourable impression of themselves. This also means that this assessment did not attempt to predict how often, or in what circumstances, a behaviour or behavioural pattern, or a shift in behavioural patterns, is likely to occur. In this way, results evenly compared behavioural traits (temperament) and their changes evenly across all participants.
Data was analysed to assess temperaments and temperament change to verify and quantify predicted changes. 231 participants were involved: 145 blue collar employees, 76 white collar employees and 10 graduates. Most white-collar employees were employed at senior management or leadership levels, and most blue-collar employees were involved in transport, manufacturing and agriculture. Due to small sample size, graduate data viewed alone is seen as unrepresentative of that demographic and so has not been shown in isolation. More participants were male than female, and more were job applicants (currently employed or unemployed) than employees not seeking to change employer. No meaningful data distinctions were found between these sub-groups. For these reasons data is only presented by blue collar, white collar, and all-group categories.
Positive (or unstressed) behaviours, are those that are generally seen as desirable socially and productively (such as patience, cooperativeness, drive, accuracy, compassion etc). Negative (or stressed) behaviours are those that are generally seen as undesirable (such as withdrawal, unproductivity, bullying, obstinance, arguing etc).
Average Temperaments – Average temperaments and shifts across all groups are shown on the chart in figure 4. Positive habits are those associated with not being under pressure, whereas negative habits are those more likely to be associated with being in a (di)stressed state. Note that temperament names have been updated for easier comprehension – choleric has become “driven, sanguine has become “exciting”, melancholy has become “detailed”, and phlegmatic has become “peaceful”.
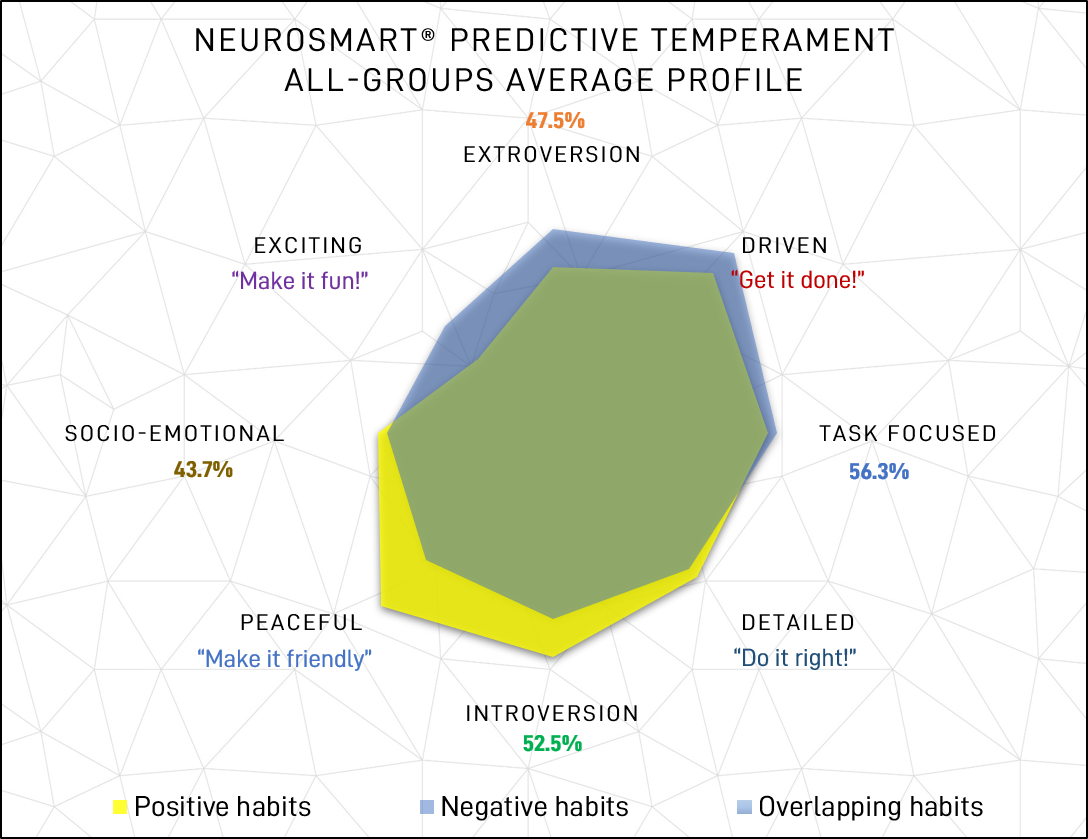 Figure 4: Plotted All-group assessment average
Figure 4: Plotted All-group assessment average
Average shift sizes are detailed in Table 1, below. Variations are calculated as a proportion of the group total rather than as a proportion of the domain level (detailed in later tables and figures).
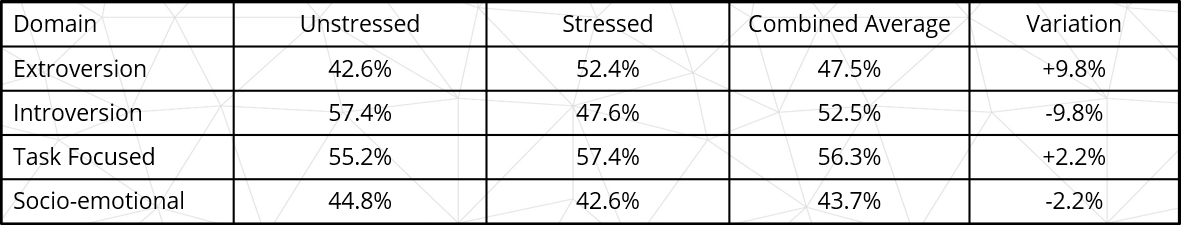 Table 1: Average shift sizes – all groups
Table 1: Average shift sizes – all groups
Blue collar and white collar role participants followed similar trends, but started from quite different bases, as shown in Table 2 below. Graduate scores, not shown separately due to the potential to mislead as a small sample size of only 10, are however included in combined averages and variation.
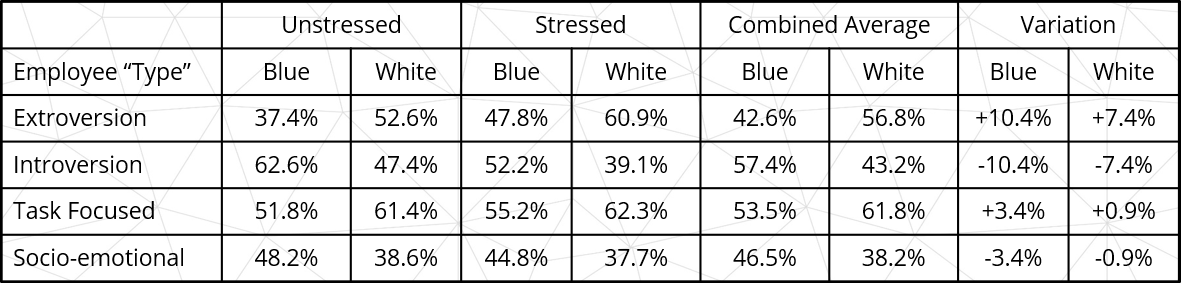 Table 2: Average shift sizes – blue- and white-collar roles
Table 2: Average shift sizes – blue- and white-collar roles
Extroversion and Introversion – The group total average profile was 42.6% extroverted and 57.4% introverted in positive (unstressed) responses, changing to 52.4% extroverted and 47.6% introverted in negative (stressed) responses. This represents an average 17.0% decrease in introversion and 22.9% increase in extroversion under stress compared to base unstressed levels.
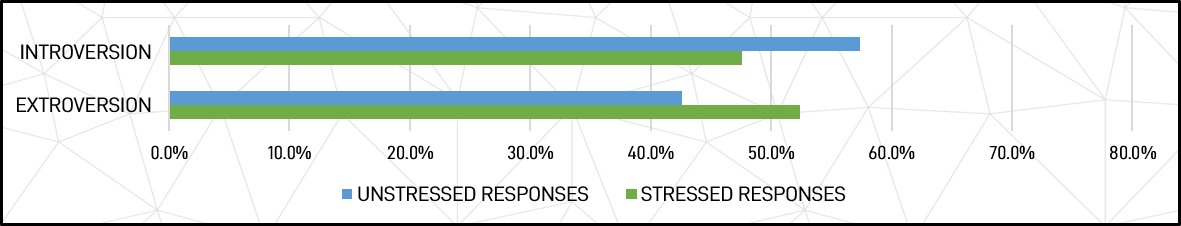 Figure 5: Introversion and Extroversion comparisons for unstressed and stressed responses
Figure 5: Introversion and Extroversion comparisons for unstressed and stressed responses
These are significant changes in behavioural patterns. Further, although there was a significant average shift from introversion to extroversion, many participants shifted the other way, as shown in Figures 6 and 7. Measured as 1 standard deviation of absolute shift values, the average shift between unstressed to stressed-state behaviours in individuals was 27.6% in introversion and 43.4% in extroversion.
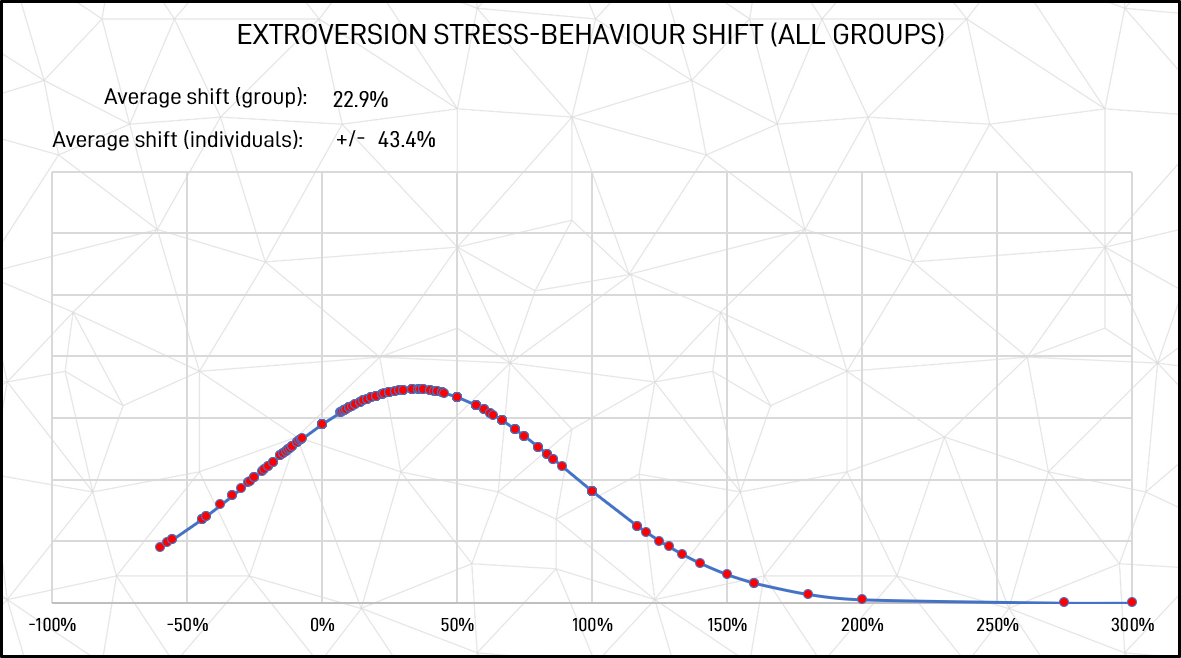 Figure 6: Extroversion stressed response shift (all groups)
Figure 6: Extroversion stressed response shift (all groups)
 Table 3: Extroversion by group
Table 3: Extroversion by group
(It is worthwhile noting that although extroversion and introversion are opposites and any gain or loss in one must be balanced by an equal loss of gain in the other, the bell-curve charts do not show opposite patterns as they originate from unequal base figures. For example, introversion starts from a higher base (unstressed response score) so, as a proportion, it’s shift to extroversion is smaller than the proportionate increase gained by extroversion for stressed response scores.)
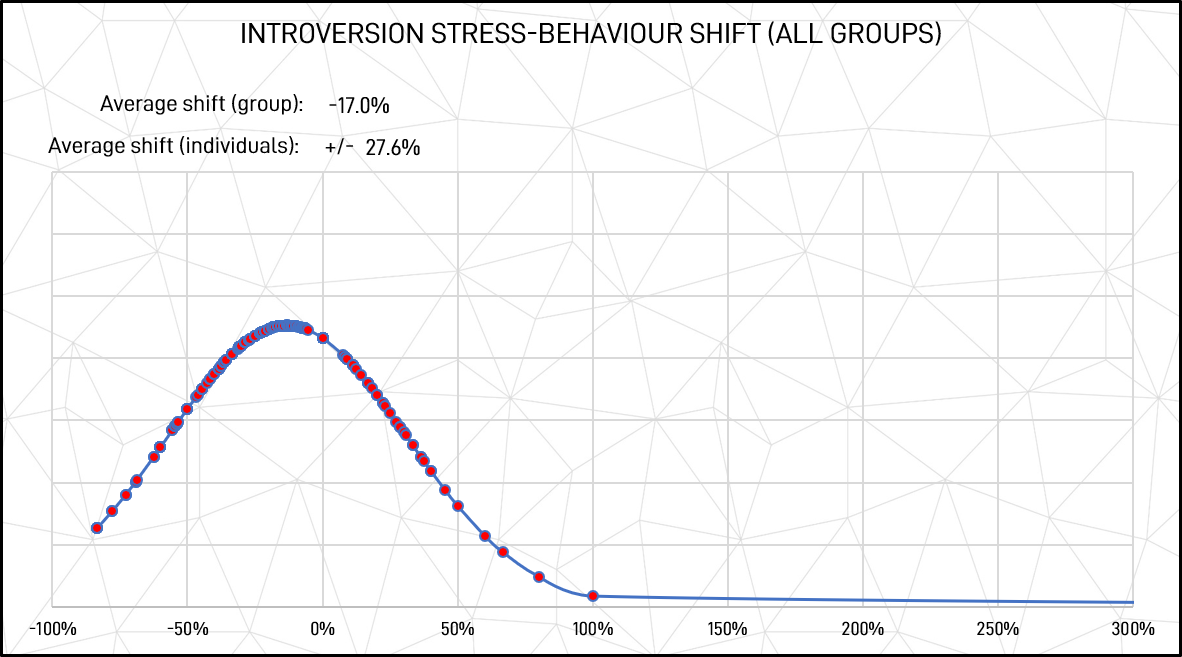 Figure 7: Introversion stressed response shift (all groups)
Figure 7: Introversion stressed response shift (all groups)
Group breakdowns show significant variations between blue- and white-collar unstressed introversion to extroversion comparisons, with both groups showing significant and similar shifts to extroversion under stress.
 Table 4: Introversion by group
Table 4: Introversion by group
The shifts of individual participants between introversion and extroversion across all groups (scored out of a possible maximum of 10) are illustrated graphically in figure 8 below.
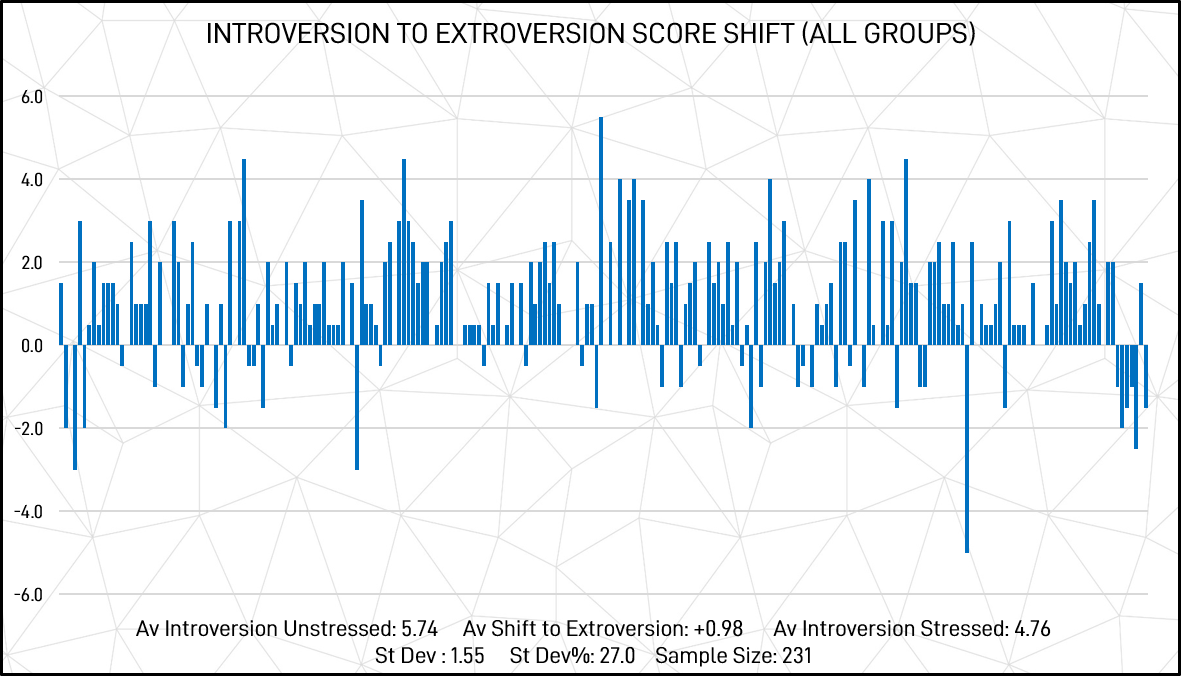 Figure 8: Introversion to extroversion score shift – individual responses
Figure 8: Introversion to extroversion score shift – individual responses
With the standard variation applied, extroversion / introversion trait variation averages for individuals can be graphically illustrated as shown in figure 8 below. Extroversion is used as the base.
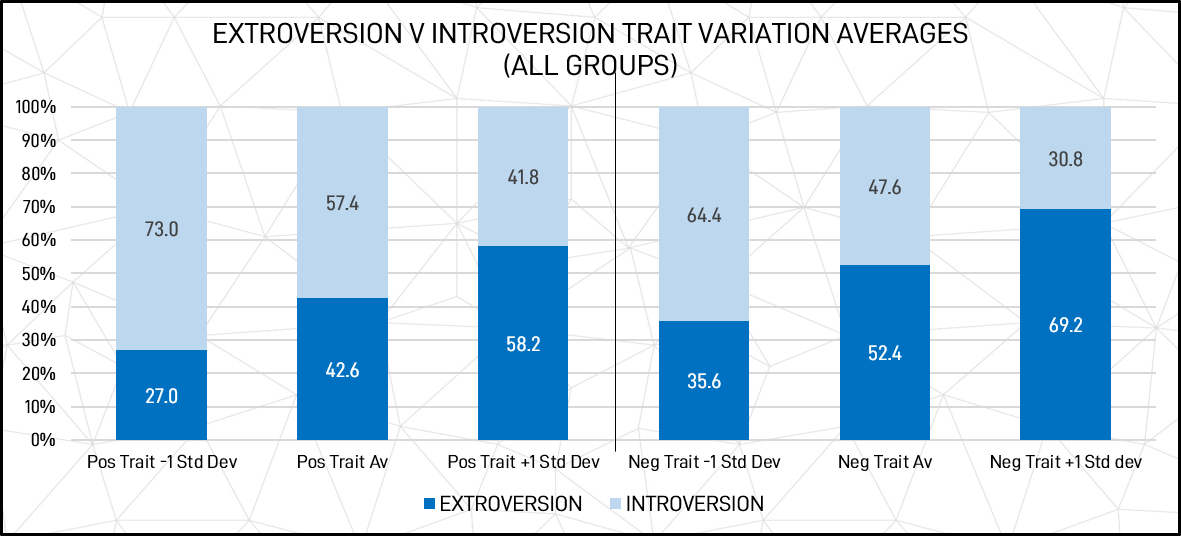 Figure 9: Introversion vs extroversion trait variation averages (1 x STDEV) for individuals
Figure 9: Introversion vs extroversion trait variation averages (1 x STDEV) for individuals
Socio-Emotional and Task Focus – The group total average profile was 44.8% socio-emotional and 55.2% task-focused in positive (unstressed) responses, changing to 42.6% socio-emotional and 57.4% task-focused in negative (stressed) responses. This represents and average 4.9% decrease in socio-emotional behaviours and a 4.0% increase in task-focused behaviours under stress compared to base unstressed levels for each.
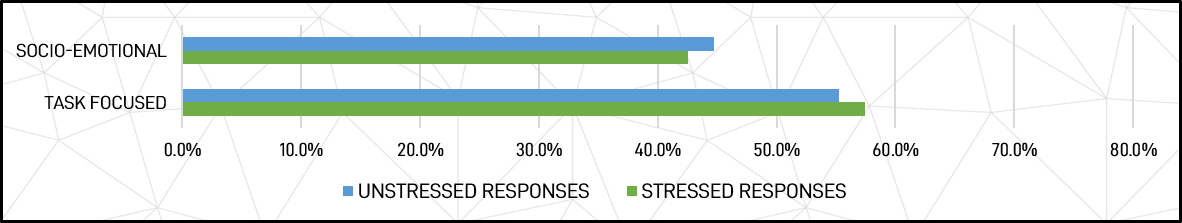 Figure 10: Socio-emotional and task focus comparisons for unstressed and stressed responses
Figure 10: Socio-emotional and task focus comparisons for unstressed and stressed responses
Group breakdowns show significant variations between blue- and white-collar unstressed socio-emotional to task focus comparisons, with both groups showing small average shifts, but significant individual shifts to task focus under stress.
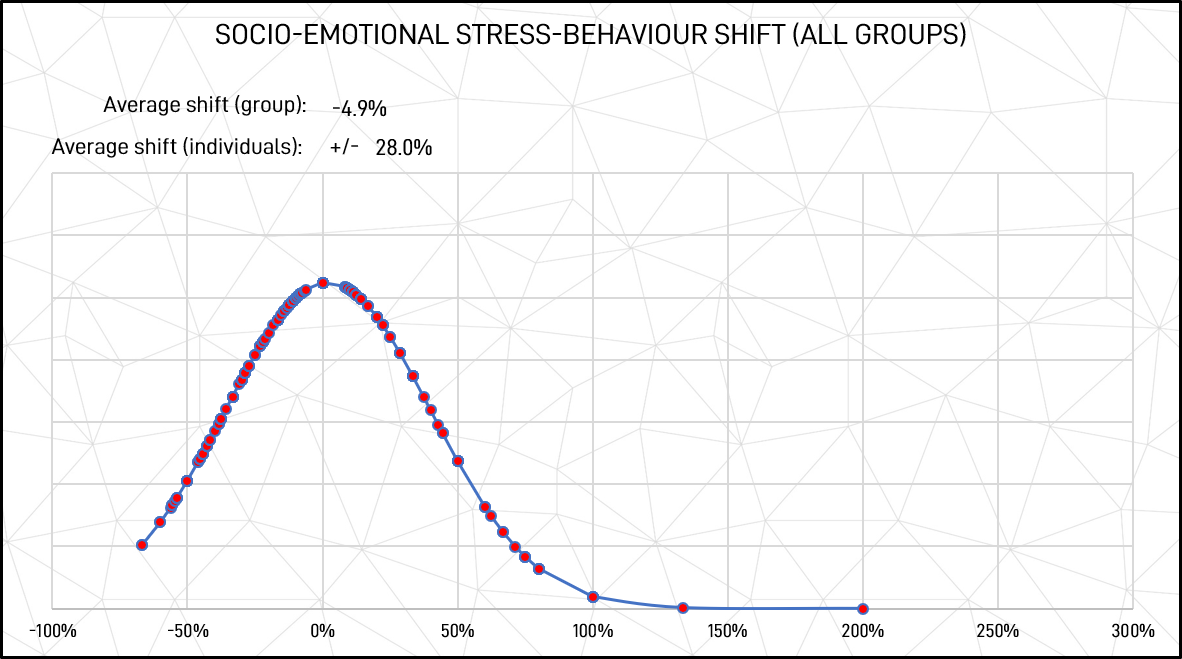 Figure 11: Socio-emotional stressed response shift (all groups)
Figure 11: Socio-emotional stressed response shift (all groups)
 Table 5: Socio-emotional responses by group
Table 5: Socio-emotional responses by group
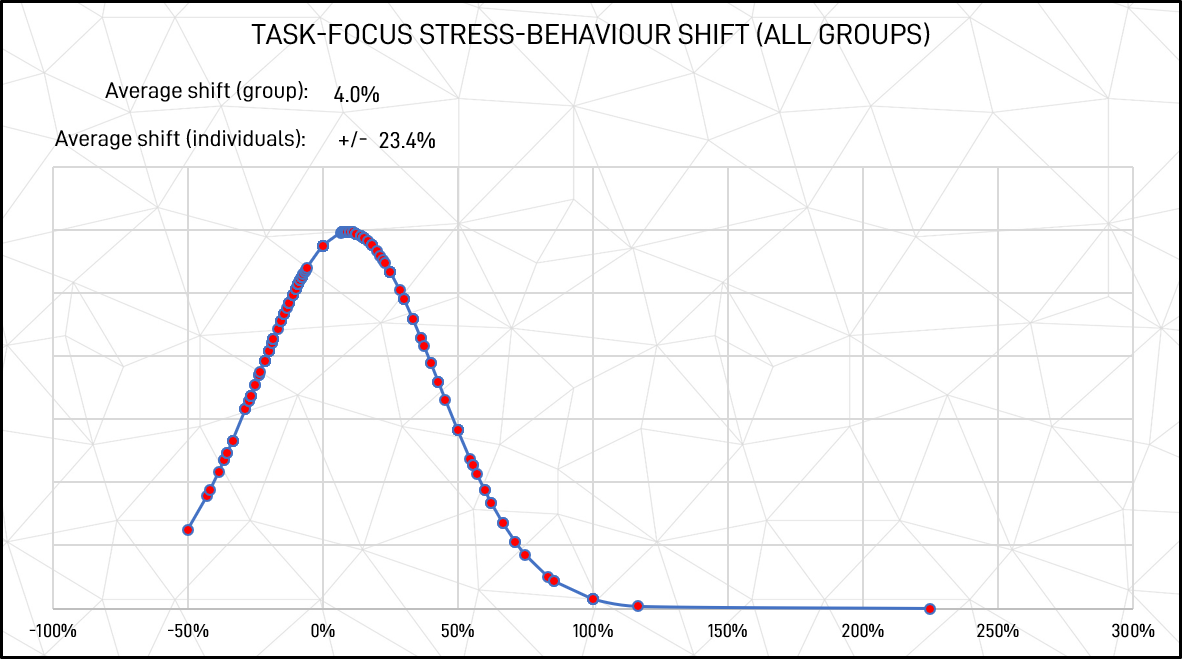 Figure 12: Task focus stressed response shift (all groups)
Figure 12: Task focus stressed response shift (all groups)
 Table 6: Task-focused responses by group
Table 6: Task-focused responses by group
As shown in figure 11, the average socio-emotional shift for individuals was 28.0%, indicating that change was common, and, with a low combined average shift, that it could almost just as easily be a strengthening of a behavioural pattern as a change. Similarly, the average individual shift from unstressed to stressed task-focused responses shown in figure 10 was 23.4%. This indicates that although the general pattern of generic behavioural stability in socio-emotional to task focus seems high, individually this is not the case.
The shifts of individual participants between socio-emotional and task focus across all groups (scored out of a possible maximum of 10) are illustrated in figure 13 below.
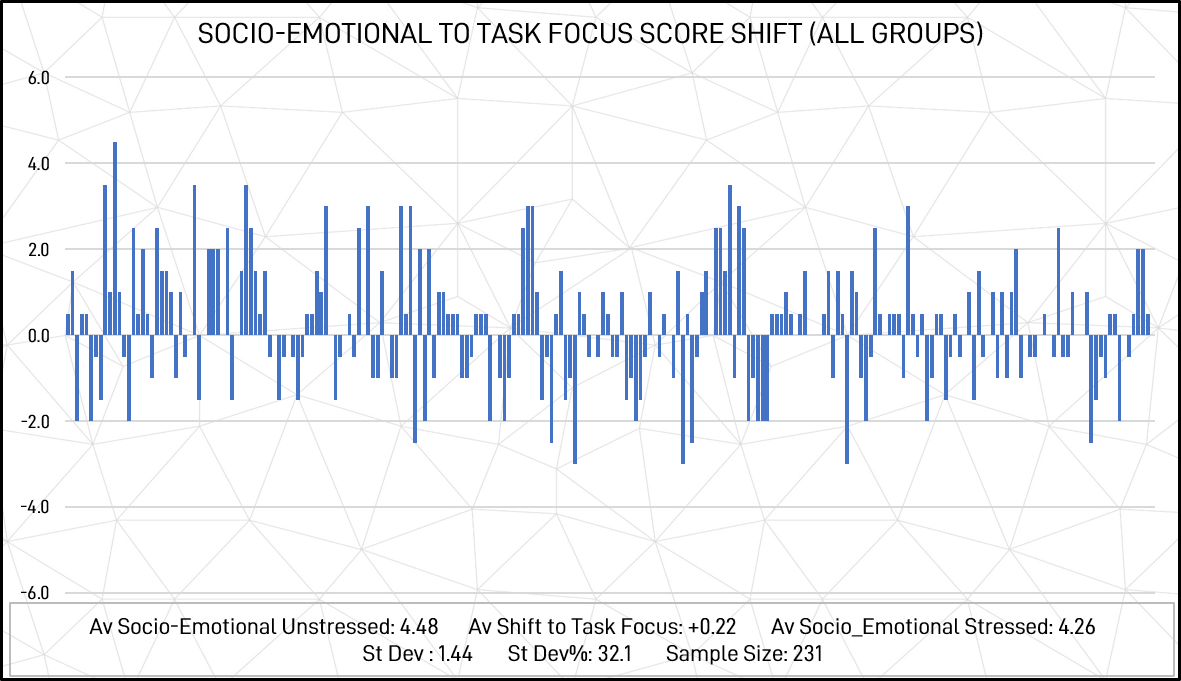 Figure 13: Socio-emotional to task focus score shifts – Individual responses
Figure 13: Socio-emotional to task focus score shifts – Individual responses
With the standard variation applied, extroversion / introversion trait variation averages for individuals are shown in figure 14 below.
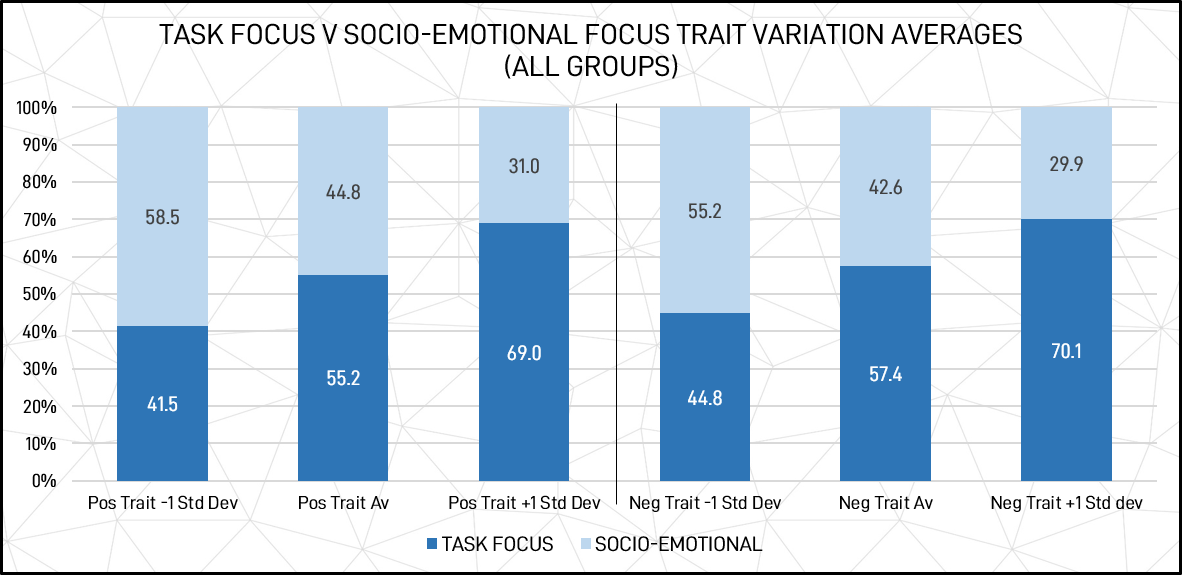 Figure 14: Socio-emotional vs task-focused trait variation averages for individuals (1 standard deviation)
Figure 14: Socio-emotional vs task-focused trait variation averages for individuals (1 standard deviation)
Summary of findings – The hypothesis that temperaments, expressed as complementary behavioural patterns, would change due to the affect of neurobiological change under stress is strongly supported by the individual participant results. Most notably, the shift from introversion to extroversion, almost certainly linked to fight over flight protective responses under high stress, was strong. There was also a shift for some individuals from extroversion to introversion, creating a high variance across the dichotomy. Similarly, although average trait variations between socio-emotional and task-focus were low, variations for individual were high, and could go in either direction. These findings indicate that behavioural shift is likely and significant under stress in potentially unpredictable ways, with single-mode or single-finding temperament assessment, categorisation or assumptions unlikely to capture the potential for any individual in the direction or extent of behavioural shift under (di)stress.
During the course of this exploration, other findings became apparent, including:
- Employees in white collar roles were significantly higher in extroversion than employees in blue collar roles when not under stress (52.6% vs 37.4%) and when under stress (60.9% vs 47.8%).
- Employees in white collar roles were higher in task focus than socio-emotional orientation than employees in blue collar roles when not under stress (61.4% vs 51.8%) and when under stress (62.3% and 55.2%).
- The impacts of behaviour pattern change due to stress were slightly but not significantly different as an average for white- and blue- collar roles (27.7% vs 31.9%).
- While some shifts were seen of greater magnitude in either direction, the vast majority individuals described shifts of less than 50% but very few indicated shifts between -2% and +2%, indicating that no and minimal change, or drastic change, are rare.
- The pattern of typology variance also showed that a minority of people, typically white collar employees in management roles in medium to large businesses, switched between opposites in drive and assertiveness when NOT under stress
Implications and observations
The four temperaments proposed by Galen and referred to under other guises by others today can be linked to the neuroscience of motivation relating to social motivation (introversion vs extroversion) on one dimension, by opposing neural networks on the other (as they relate to socio-emotional and task-specific focus of attention), and by genetics, brain development, neuroplasticity and neuro-stability through decreased potential for change with maturity, fundamentally.
That other assessments have not explored the cause behind their observations or dealt with the implications of neuroplasticity and neurobiology in trait formation and change hasn’t stopped them from being reasonably reliable as a generalist approach much of the time. However, the neurobiology of stress (FFF responses, sympathetic and parasympathetic nervous system dominance, learned behavioural habit change linked to synaptogenesis and myelinogenesis relating to those systems) has been generally ignored, severely limiting their reliability in predicting behaviours in differing contexts and environments.
For this reason, neurobiological stress response undermines the accuracy and usefulness of previous fixed-temperament ideas. Rather, neurobiology predicts, and our research supports, the hypothesis that while behaviour under pressure can occur in ways that are reasonably consistent with a given profile with only minor variations, others show significant changes, often switching to behavioural responses that don’t fit a general temperament profile, and that those changes can go in directions not predicted by single-mode or single-point temperament assessments.
The extent and nature of stress-related temperament change, while reasonably likely, is variable and cannot be presumed to be reliably predictable from a simplistic assessment or assumption based on behaviours described or witnessed in a single scenario. Our research has shown that while it is unlikely that an individual will give up most of the behaviours associated with a dominant temperament as part of their stress response, it is quite common that stress response changes are significant enough to see them behaving in ways that might be otherwise unexpected. Further, where participants score low in one domain, it is almost certain that their stressed responses in that domain wil increase or decrease markedly. The absolute stability inferred by other assessments, occurring in less than 1% of cases, might be described as the statistical outlier.
Shifts toward extroversion are more common than shifts toward introversion, indicating that more people, when under extreme stress, are likely to exhibit behaviours in pro-actively “defending themselves or others” (fight response) than of retreating and avoiding (flight response). Similarly, there is also a small average tendency toward task focus when under stress, which may relate to “factual” rather than “emotional” resourcing in conflict despite limited capacity under stress to do so objectively.
There is another danger in simplification of assessment scores not described by average statistics. The pattern of variance showed a minority of white-collar employees changing between opposites in drive and assertiveness when NOT under stress, perhaps as learned adaptive responses to subtle changes in power-sensitive contexts, either in work or in earlier life. This would see them categorised generically as being neither assertive or submissive, missing the essence of their temperament. However, this “political savvy”, in face to face job interviews, has not gone unnoticed by employers in the private sector that we have worked with (without prior reference to the assessment), leading them to express “not getting” or “not trusting” an individual, as predicted by Fornito et al (30).
In our research, this power-sensitive temperament has been more common for senior managers in large organisations, but rarely for others in those organisations or for similar roles in private and smaller businesses, suggesting sensitivity and moderation in exercising power and influence where political factors are more likely to contribute to survival and career success than in smaller or more directly outcome-accountable roles. This presents a serious credibility and applicability problem for simplistic temperament assessments, where opposites simply cancel each other out, resulting in a meaningless graphical mid-point that fails to capture any of that important nuance.
It might also be speculated that, as with any diagnosis, individuals resonate with descriptions of themselves made by themselves, irrespective of contrary factors. For this reason, it is easy to find areas of agreement with temperament assessments, especially ones that allow for a high proportion of positive traits, just as fortune-tellers and other frauds routinely do to gain credibility. In this way, there is a danger in any assessment, especially where it is simply a re-encapsulation of what a participant has already identified with, as being little more than cold-reading, confirmation and validation. The NEURO-M Predictive Temperament Assessment, by not allowing a preferred picture, and by acknowledging neurobiological change under stress, limits such popularist validation through preferred identity confirmation.
The differences between blue- and white-collar employees in base trait level were meaningful. However, no inference was made as to whether those differences result from behavioural responses to differing role demands or whether those roles simply attract different temperament blends. In other words, this correlation could just as easily be a cause as a result.
It was notable that temperament stability, and the stress-related behavioural stability that underpins it, was only slightly higher for white-collar employees than blue-collar employees. This indicates that despite seniority, emotional and behavioural self-management for managers and other white collar employees cannot be assumed to be superior to blue collar employees, strongly suggesting that managers are not more highly skilled in emotional intelligence, despite strong links between EQ and leadership effectiveness (33) and, presumably, some forms of management training for at least some of them.
It is reasonable to expect that environments, both physical and psychological, that are perceived to be unsafe are likely to trigger behavioural shifts more often and/or to a greater extent than those perceived to be safe. Similarly, it is reasonable to expect that individuals with greater amygdala sensitivity either generally (anxiety-related) or specifically (contextually-related) are likely to shift to stress responses more frequently and/or severely. It is also likely that non-work stressors will influence stress tolerance and response intensity in the workplace.
Limitations
Despite differences in assessment base and philosophy, information nuance and predictive usefulness, temperament assessments typically don’t attempt precision in predicting behaviours in specific situations or the stress levels at which behaviours will occur. Similarly, the NEURO-M Predictive Temperament Assessment does not attempt to judge thresholds, contextual triggers or frequency of behaviour change. Other forms of assessment (e.g., the NEURO-M Motivation Assessment) and behavioural interviews are more effectively useful for exploring such contextually-relevant tolerances.
As with any form of self-assessment, there are limitations on the capacity for any individual for self-objectivity in completing temperament assessments, along the potential for participants to present an overtly preferred, memory-selective or mood-affected picture of themselves, despite mechanisms that make this more difficult. There are also opportunities for participants to interpret questions or words differently than intended, or to simply make mistakes when answering. Despite this, overall reliability of well-designed assessments appears high, at least to the extent of the philosophy or rationality they employ and the limitations of their usefulness.
Further, with careful design, along with other forms of assessment and interviews, the likelihood of these types of errors and manipulations can be substantially reduced or detected. Considering the morality of providing any type of judgemental information, and, in a practical sense, its frequent use in recruitment and career planning in an environment where the credibility of the assessment service provider was closely linked to long-term result accuracy, this approach was taken with the NEURO-M Predictive Temperament Assessment in its preferred role as a part of an overall process, to maximise reliability, irrespective of the intentions of participants.
Conclusion
Brain growth and development ensure that particular responses will be more favoured by individuals than others, influenced by DNA, life experience, context, environment and other factors. The repetition of these behavioural patterns are known as traits, and the combination of these traits is described as a temperament.
Popular temperament assessments plot behavioural patterns to sense commonalities, or traits. They use the categorisation of trait patterns to describe common typologies and combinations, with predictable generic responses, a proposal that is supported by synaptogenesis and myelinogenesis.
However, it is apparent that a single, generic temperament typology is not supported by the sympathetic nervous system’s stress response mechanism, most notably the FFF response system. Rather, it is more likely that individuals have a non-stressed temperament and a stressed temperament, the variance being in the extent of stress experienced and the nature of habitual (myelinated) responses in both conditions.
The NEURO-M Predictive Temperament Assessment results show that significant behavioural differences occur that sit outside of otherwise predictable patterns under (di)stress. These changes are quantified as significant changes both in introversion vs extroversion and socio-emotional vs task-focused behaviours on the other for individuals.
This means that:
- The concept of stable single-temperaments under stress is not supported
- There is a general, but highly variable and generically unpredictable (by other popular assessments), shift toward extroversion, likely expressed and/or interpreted as assertiveness or aggression, under stress.
- There is a smaller, but variable and generically unpredictable, shift toward task-focused behaviours, likely expressed as narrowing of focus around specific “facts”, despite reduced capacity to do so objectively and openly.
- Behavioural change can occur in any direction, with a reasonably strong shift toward extroversion from introversion, and a weaker shift from socio-emotional orientation toward task focus.
- Assessments that do not distinguish between stressed and non-stressed behavioural response patterns as separate “sub-temperaments”, and/or that attempt to over-simplify results in any situation or context, are unlikely to be usefully accurate or meaningful.
For these reasons, and to the extent outlined by the theory and research detailed here, the NEURO-M Predictive Temperament Assessment, and the hypothesis that it is more accurate than non-neurologically-based alternatives, is both validated and supported.
References:
- Kushner I. The acute phase reactants. In: Kelley WEH, Ruddy S, Sledge C, editors. Textbook of rheumatology. 2nd ed. Philadelphia PA: W. B. Saunders; 1985. p. 653-64.
- Fähræus R. The suspension stability of the blood. Physiological revolutions. 1929;9(2):241-74.
- Kushner I. The 4 humors enthrocyte sedimentation: The most influential observation in history. American journal of medical sciences. 2012;346(2):154-7.
- Kagan J. Galen’s Prophecy: Temperament in human nature. New York, NY: Basic Books; 1998.
- Merenda PF. Toward a four-factor theory of temperament and/or personality. Journal of personality assessment. 1987;51:367-74.
- Spinath F. Genetic and environmental influences on observed personality: Evidence. Journal of personality and social psychology. 2001;80(4):655-68.
- Bleidorn W. Patterns and sources of adult personality development: Growth. Journal of personality and social psychology. 2009;97(1):142-55.
- Carmichael CM. A longitudinal family study personality change and stability. Journal of personality. 1994;62(1):1-20.
- Siegel DJ. The pocket guide to interpersonal neurobiology: An integrative handbook of the mind. New York, NY: Norton; 2012.
- Kawamoto T, Endo T. Genetic and environmental contributions to personality trait stability and change across adolescence: Results from a Japanese twin sample. Twin research and human genetics. 2015;18(5):545-56.
- Hopwood CJ, Donnellan MB, Blonigen DM, Krueger RF, Mcgue M, Iacono WG, et al. Genetic and environmental influences on personality trait stability and growth during the transition to adulthood: A three-wave longitudinal study. Journal of personality and social psychology. 2011;100(3):545-56.
- Soto CJ, John OP, Gosling SD, Potter J. Age differences in personality traits from 10 to 65: Big Five domains and facets in a large cross-sectional sample. Journal of personality and social psychology. 2011;100:330-48.
- Arnett JJ. Emerging childhood: A theory of development from the late teens through the twenties. American Psychologist. 2000;55:469-80.
- Gallagher P, Fleeson W, Hoyle RH. A self-regulatory mechanism for personlaity trait stability: Contra-trait effort. Social psychological and personality science. 2011;2(4):335-42.
- Lewis GJ, Cox SR, Booth T, Muñoz Maniega S, Royle NA, Valdés Hernández M, et al. Trait conscientiousness and the personality meta-trait stability are associated with regional white matter microstructure. Social cognitive and affective neuroscience. 2016;11(8):1255-61.
- Hampson SE, Goldberg LR. A first large cohort study of personality trait stability over the 40 years between elementary school and midlife. Journal of personality and social psychology. 2006;91(4):763-79.
- Grawe K. Neuropsychotherapy. New York: Psychology Press; 2007.
- Craig A, Silk T. Schedules of reinforcement, learning, and frequency reward programs. Advances in Consumer Research. 2009;36:555-6.
- Rosenau PM. Post-modernism and the social sciences: Insights, inroads, and intrusions. Princeton, NJ: Princeton university press; 1992.
- Heider F. The Psychology of Interpersonal Relations. Hillsdale, NJ: Lawrence Erlbaum Associates; 1958.
- Weiner B. Attribution Theory. In: O’Connor T, Sandis C, editors. A companion to the philosophy of action. Chichester, UK: Wiley-Blackwell; 2010.
- Rossouw PJ, editor. Neuropsychotherapy: Theoretical underpinnings and clinical applications. St Lucia, AU: Mediros; 2014.
- Potts GF. Impact of reward and punishment motivation on behavior monitoring as indexed by the error-related negativity. International journal of psychophysiology. 2011;81(3):324-31.
- Kim SI. Neuroscientific model of motivational process. Frontiers in Psychology. 2013;4.
- Rangel A, Camerer C, Montague PR. A framework for studying the neurobiology of value-based decision-making. Nature reviews neuroscience. 2008;9:545-56.
- Boyatzis RE, Rochford K, Jack AI. Antagonistic neural networks underlying differentiated leadership roles. Frontiers in human neuroscience. 2014;8(Article 114):1-15.
- Csikszentmihalyi M. Flow: The psychology of optimal experience. New York, NY: HarperCollins; 1991.
- Jack AI, Dawson A, Begany K, Leckie RL, Barry K, Ciccia A. fMRI reveals reciprocal inhibition between social and physical cognitive domains. Neuroimage. 2012;66C:385-401.
- Jack AI, Dawson A, Norr M. Seeing human: distinct and overlapping neural signatures associated with two forms of of dehumanisation. Neuroimage. 2013;79:313-28.
- Fornito A, Harrison BJ, Zalesky A, Simons JS. Competitive and cooperative dynamics of large scale brain networks supporting recollection. Proceedings of the National Academy of Sciences of the USA. 2012;109:12788-93.
- Beissner F, Meissner K, Bat K, Napadow V. The autonomic brain: An activation likelihood meta-analysis for central processing of autonomic function. Journal of neuroscience. 2013;33:10503-11.
- Fox MD, Snyder AZ, Vincent JL, Corbetta M, van Essen DC, Raichle ME. The human brain is intrinsically organised into dynamic, anticorrelated functional networks. Proceedings of the national academy of sciences of the United States of America. 2005;102:9673-8.
- Goleman D. Emotional Intelligence. New York: Bantam House; 2006.